ABSTRACT
The prevalence of metabolic syndrome caused by diets containing excessive fatty acids is increasing worldwide. Patients with metabolic syndrome exhibit abnormal lipid profiles, chronic inflammation, increased levels of saturated fatty acids, impaired insulin sensitivity, excessive fat accumulation, and neuropathological issues such as memory deficits. In particular, palmitic acid (PA) in saturated fatty acids aggravates inflammation, insulin resistance, impaired glucose tolerance, and synaptic failure. Recently, adiponectin, brain-derived neurotrophic factor (BDNF), and glucose-like peptide-1 (GLP-1) have been investigated to find therapeutic solutions for metabolic syndrome, with findings suggesting that they are involved in insulin sensitivity, enhanced lipid profiles, increased neuronal survival, and improved synaptic plasticity. We investigated the effects of adiponectin, BDNF, and GLP-1 on neurite outgrowth, length, and complexity in PA–treated primary cortical neurons using Sholl analysis. Our findings demonstrate the therapeutic potential of adiponectin, BDNF, and GLP-1 in enhancing synaptic plasticity within brains affected by metabolic imbalance. We underscore the need for additional research into the mechanisms by which adiponectin, BDNF, and GLP-1 influence neural complexity in brains with metabolic imbalances.
-
Keywords: Metabolic syndrome; Adiponectin; BDNF; GLP-1; Palmitic acid
INTRODUCTION
The prevalence of metabolic syndrome accompanying obesity is increasing dramatically worldwide [
1]. Diets containing excessive saturated fatty acids, such as palmitic acid (PA), accelerate the development of metabolic complications, including obesity, diabetes mellitus, glucose intolerance, inflammation, and insulin resistance [
2]. PA has been shown to induce impaired glucose metabolism, inflammatory responses [
3], insulin sensitivity attenuation [
4], and the autophagic flux pathway in neurons [
5]. A recent study found that PA could aggravate memory loss and neuroinflammation by stimulating the production of pro-inflammatory cytokines, such as tumor necrosis factor alpha [
6], through the nuclear factor kappa-light-chain-enhancer of activated B cells signaling pathway [
7]. High levels of PA promote cell death, impaired neural differentiation, mitochondrial dysfunction, and abnormal neurite outgrowth [
8]. To overcome the synaptic dysfunction observed in people with obesity, it is essential to study neuronal dysfunction under PA toxicity and identify drugs that can improve it.
Adiponectin is a 30-kDa hormone produced by adipose tissue that is found at lower levels in patients with obesity and diabetes [
9]. Adiponectin promotes insulin sensitivity, regulation of fatty acid metabolism, antioxidant responses, and anti-inflammatory responses [
10]. Adiponectin contributes to synaptic plasticity, ATP production rates, glucose uptake, neuronal excitability, neuronal survival, neurogenesis, and glial cell activation in the central nervous system [
11]. In patients with Alzheimer’s disease (AD), a reduction in adiponectin levels is observed in the blood, accompanied by synaptic failure [
12].
Brain-derived neurotrophic factor (BDNF), a member of the neurotrophin family, promotes axonal branching, synapse refinement, dendritic growth, synapse maturation, induction of long-term potentiation (LTP), and spine maturation [
13]. A lower level of circulating BDNF is observed in the sera of patients with metabolic syndrome, obesity [
14], and psychiatric disease [
15].
Glucagon-like peptide-1 (GLP-1), a glycoprotein hormone produced by gut L-cells and specific neurons in the nucleus tractus solitarius of the brain [
16], improves impaired glucose metabolism, reduces the sensation of hunger, and reduces food intake in patients with metabolic syndrome and obesity [
17]. Lower GLP-1 levels have been associated with obesity [
18]. GLP-1 promotes neuronal survival and neurite outgrowth in PC12 neurons [
19] by modulating cyclic adenosine monophosphate (cAMP) signaling [
20]. GLP-1 also regulates synaptic transmission, neuronal function [
21], calcium channel–related signaling [
22], depolarization of neuronal cell membrane potential, and spike firing rates [
23]. GLP-1 modulates neuronal activity through signaling pathways such as the mitogen-activated protein kinase extracellular signal–regulated kinases 1 and 2, protein kinase C, and protein kinase A (PKA) pathways [
22]. Additionally, GLP-1 can inhibit neuroinflammation and enhance energy metabolism, neural connectivity, and neural structure [
24] in models of neurodegenerative diseases such as AD [
25].
As mentioned, adiponectin, BDNF, and GLP-1 are known to influence neuronal survival, synaptic plasticity, neural structure, and neuronal activity in neurons under conditions of metabolic imbalance. However, there have been no studies on the effects of adiponectin, BDNF, and GLP-1 on neural structure in PA-treated neurons. Thus, using Sholl analysis, we investigated the effects of a 30-kDa adipocyte complement–related protein (Acrp30—as an adiponectin globular form), BDNF, and exendin-4 (as a GLP-1 analog) on neuronal structure and neurite outgrowth in PA-treated cortical neurons.
MATERIALS AND METHODS
Mouse primary cortical neuron (PCN) culture
PCNs were isolated from the cerebral cortices of C57BL/6 mouse embryos provided by Koatech (Pyeongtaek, Korea) on embryonic day 14. Cortices were isolated and then incubated in 0.25% trypsin containing dissection/dissociation medium (1X Hank’s balanced salt solution, 20% glucose solution, 1X sodium pyruvate, and 1 M HEPES [4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid] were purchased from Thermo Scientific, Waltham, MA, USA) for 15 min with inverting every 5 minutes. Cells were triturated to single cells in a plating medium (Minimum Essential Medium containing Earle’s balanced salt solution, 10% fetal bovine serum, 20% glucose solution, 1X sodium pyruvate, 1X GlutaMAX, and 100 U/mL penicillin-streptomycin) and incubated for 2 hours in 5% CO2 at 37°C. The plating medium was replaced with a maintenance medium (Neurobasal medium, 1X B27, 1X GlutaMAX, 100 U/mL penicillin-streptomycin) for cell growth. The maintenance medium was replaced every 3 days.
PCNs were pretreated for 2 hours with PA (250 μM) [
26], Acrp30 (100 ng/mL) [
27], BDNF (50 ng/mL) [
28], and exendin-4 (10 nM) [
8] to analyze neurite complexity on a day in vitro 7 (DIV7).
For neurite complexity visualization, PCNs were transfected with the pMAX-GFP plasmid (Lonza, Lake Charles, LA, USA) with FuGENE 6 transfection reagent (Promega, Madison, WI, USA) for 48 hours, according to the manufacturer’s instructions.
Sholl analysis
For the neurite complexity analysis, PCNs were transfected with a green fluorescent protein (GFP) using the FuGENE 6 transfection reagent (Promega). Neurite complexity was visualized using an Eclipse Ts2 fluorescence microscope (Nikon, Tokyo, Japan). Neurite complexity was analyzed using the Sholl analysis plugin in ImageJ (National Institutes of Health, Bethesda, MD, USA). Seven PCNs were selected and analyzed in a blinded manner. With a point tool, the soma center was selected and used to quantify size-related parameters, such as total neurite length and the number of intersections, from radii of 10 μm and 700 μm, with a step size of 10 μm. Samples were set at 3 per radius, and the degree of polynomial fit was set as the “best-fitting degree.” We selected and combined data from the intersection columns of the Sholl profile list.
Statistical analysis
All data were analyzed using unpaired 2-tailed t-tests with Welch’s correction between the groups. Data were considered significant at p <0.05 and p < 0.01 in the statistical analysis.
RESULTS
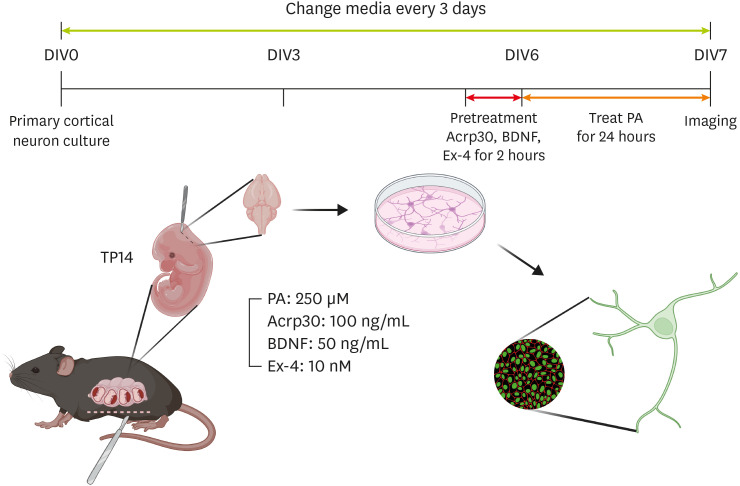
We investigated the effects of adiponectin, BDNF, and GLP-1 on neurite outgrowth in PCNs. The PCNs were treated according to a treatment plan (
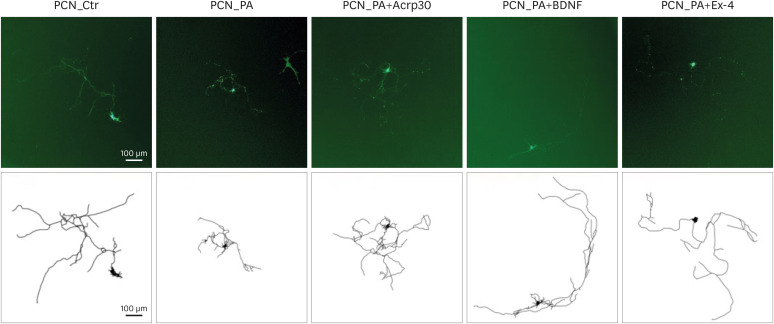
Figure 1). We performed a morphological analysis of the GFP-transfected PCN images (
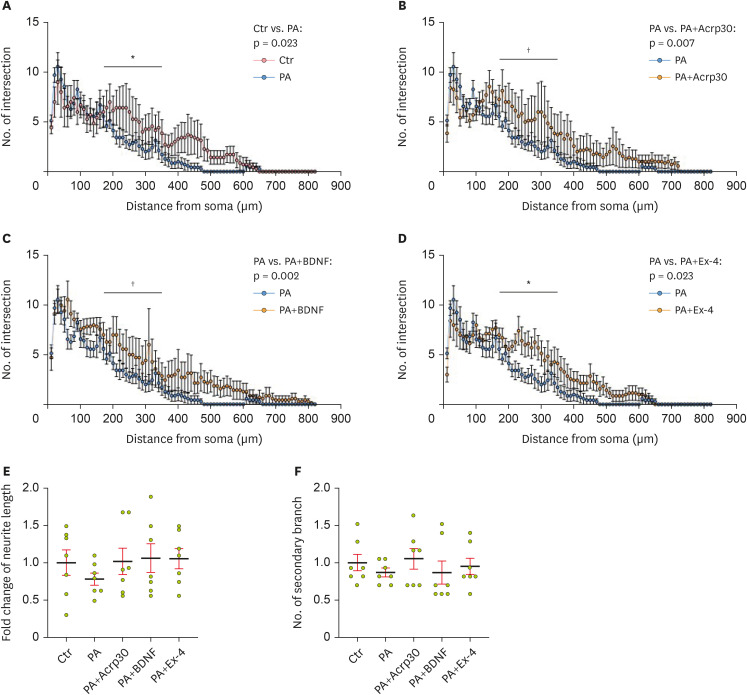
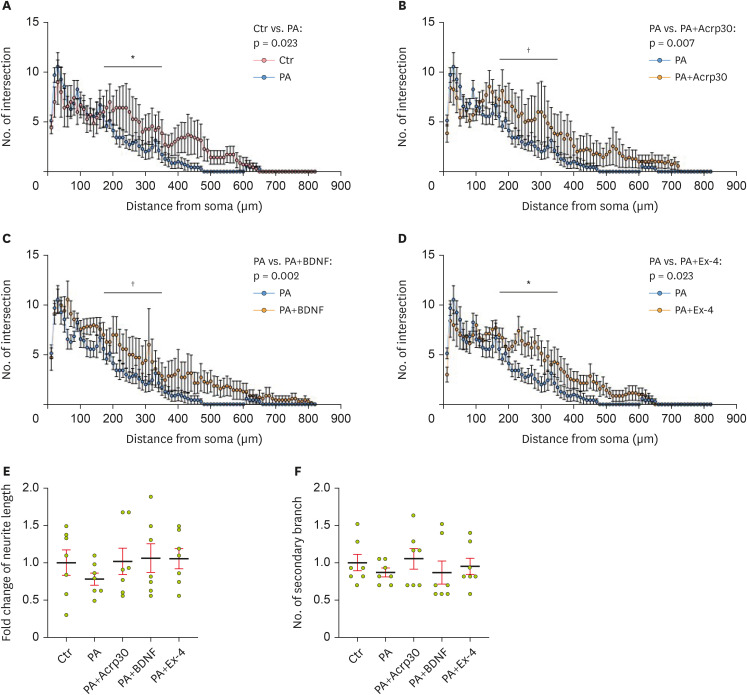
Figure 2). PCNs were analyzed using Sholl analysis and in terms of neurite complexity, encompassing the neurite length, number of neurites from the soma, and number of secondary branches (
Figure 3). The intersection of neurites from the soma was lower in PA-treated PCNs than in control neurons (
Figure 3A). However, under PA-induced metabolic imbalance conditions, the decreased intersection of neurites in PCNs was increased by pretreatment with Acrp30 (
Figure 3B). BDNF also increased the intersection of neurites in the PCNs, which was reduced by PA treatment (
Figure 3C). Additionally, exendin-4 treatment ameliorated the PA-induced reduction in neurite intersections (
Figure 3D). The total neurite length was shorter in PA-treated PCNs than in control neurons (
Figure 3E). However, Acrp30, BDNF, and exendin-4 treatment tended to increase neurite length, which was decreased by PA in PCNs, indicating that Acrp30, BDNF, and exendin-4 enhance neurite outgrowth through metabolic damage. Furthermore, the number of secondary branches, which was reduced in PA-treated PCNs, was increased by treatment with Acrp30, BDNF, and exendin-4 (
Figure 3F). Collectively, these results indicate that adiponectin, BDNF, and GLP-1 pretreatment tended to enhance neurite complexity in terms of parameters such as neurite outgrowth and neurite formation under conditions of PA-induced metabolic imbalance.
Figure 1
Schematic image of drug treatment in primary cortical neurons. This schematic shows the study’s experimental plan.
DIV, day in vitro; BDNF, brain-derived neurotrophic factor; Ex-4, exendin-4; PA, palmitic acid.
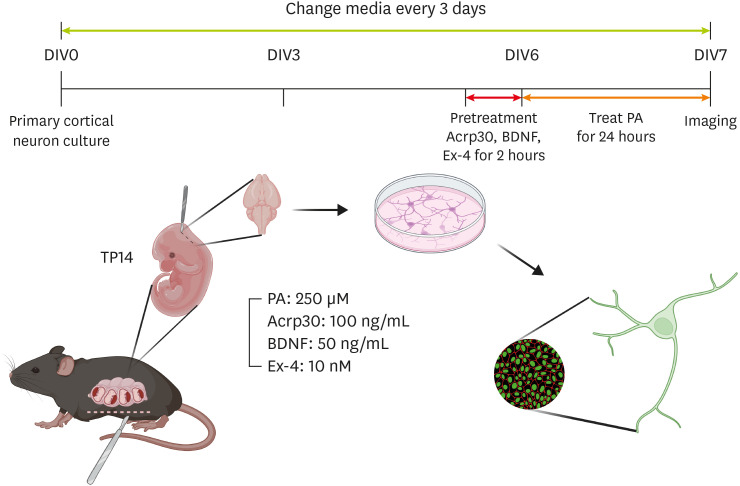
Figure 2
Fluorescence images of primary cortical neurons. Fluorescence images of neurite complexity in green fluorescent protein: transfected PCNs on day in vitro 7 after treatment with vehicle (Ctr), PA, PA+Acrp30, PA+BDNF, and PA+Ex-4. The scale bars represent 100 μm.
PCN, primary cortical neuron; PA, palmitic acid; BDNF, brain-derived neurotrophic factor; Ex-4, exendin-4.
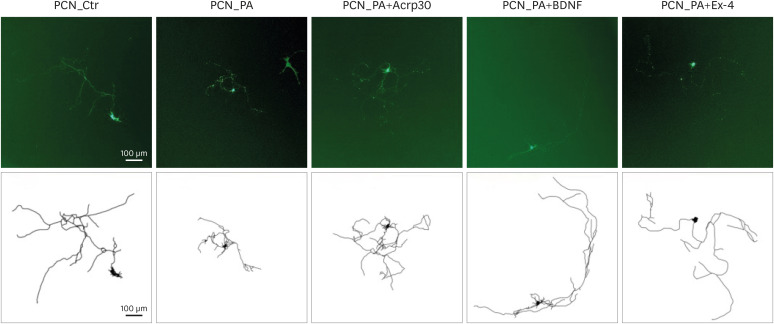
Figure 3
Sholl analysis for neurite complexity in primary cortical neurons. (A) Histogram of neurite intersection in PCNs on DIV7 after treatment with vehicle (Ctr) and PA. (B) Histogram of neurite intersection in PCNs on DIV7 after treatment with PA, as well as PA+Acrp30. (C) Histogram of neurite intersection in PCNs on DIV7 after treatment with PA, as well as PA+BDNF. (D) Histogram of neurite intersection in PCNs on DIV7 after treatment with PA, as well as PA+Ex-4. (E) Histogram of neurite length after treatment with reagents. (F) Histogram of secondary branches on DIV7 after treatment with reagents. Data information: PCNs were treated with 250 μM of PA, 100 ng/mL of Acrp30, 50 ng/mL of BDNF, and 10 nM of Ex-4 in each group according to the treatment plan. (A-D) Data are shown as the mean ± standard error of the mean.
PCN, primary cortical neuron; DIV7, day in vitro 7; PA, palmitic acid; BDNF, brain-derived neurotrophic factor; Ex-4, exendin-4.
*p < 0.05 and †p < 0.01 (unpaired 2-tail t-test with Welch’s correction).
DISCUSSION
We investigated the effects of Acrp30, BDNF, and exendin-4 on neurite outgrowth and neural structure in PA-treated cortical neurons. Our study findings suggested that excessive dietary intake of saturated fatty acids, such as that associated with a PA-enriched diet, aggravates synaptic plasticity, neuronal death, and abnormal neurite outgrowth, leading to cognitive decline [
29]. First, we identified that adiponectin improves neurite length and contributes to the complicated neural structure of PA-treated cortical neurons. Acrp30, a secreted adiponectin protein expressed in differentiated adipocytes [
30], belongs to a family of proteins containing the C1q globular domain [
31]. In a previous study, Acrp30 was shown to regulate insulin sensitivity, mitochondrial biogenesis, and inflammation [
32]. Adiponectin exerts multiple regulatory roles by binding the adiponectin receptors AdipoR1 and AdipoR2, which are expressed in brain regions that contribute to learning and memory function, such as the cerebral cortex and hippocampus [
33]. A recent study found that adiponectin could enhance neurite outgrowth, memory loss, and synaptic complexity in both AD and obesity-related cognitive impairment [
34]. Adiponectin administration, including the administration of the adiponectin homolog osmotin, has been shown to enhance LTP impairment [
35] and rescue memory loss in AD mouse models through AdipoR activation [
36]. Adiponectin mimics the drug AdipoRon and improves fear-related memory deficits and synaptic transmission [
37]. Zhang et al. [
38] showed an improvement in synaptic density, increased dendritic spine maturation, and increased neurite length through AdipoR1. According to our data, globular adiponectin from Acrp30 maintains neural complexity and enhances synaptic plasticity in obese brains with high levels of PA.
Our data showed that the mature form of BDNF improved neurite outgrowth and maintained a complicated neural structure in PA-treated cortical neurons. Some studies have shown that mature BDNF ameliorates neurite outgrowth, synaptic function, axon sprouting, and neuronal excitability in hippocampal neurons [
39]. In contrast, some studies have demonstrated that pro-form BDNF suppresses neurite outgrowth in primary cultured neurons through the activation of RhoA [
40]. According to our data and previous evidence, mature BDNF enhances neural complexity and synaptic plasticity in the brains of obese animal models with high PA levels.
Finally, our data show that exendin-4, a GLP-1 analog, enhances neurite outgrowth and maintains the neural structure of PA-treated cortical neurons. Exendin-4 is a39–amino acid peptide agonist of GLP-1 [
41] that increases insulin sensitivity in diabetes following cAMP/PKA signaling [
42]. A recent study found that exendin-4 improved memory loss, synaptic plasticity, and neuronal death in hippocampal neurons in a streptozotocin-induced AD rat model [
43]. Exenatide, a synthetic version of exendin-4, has been shown to improve cognitive performance and activate the expression of neurotrophic factors in a diabetic mouse model [
44]. Additionally, exendin-4 facilitates rearrangement of the actin cytoskeleton and influences the axon growth cone in SH-SY5Y and PC12 neurons by downregulating RhoA activation [
45]. Exendin-4 improves neurite dendritic growth, spine maturation, and LTP induction in high-fat diet-fed mice and in vitro primary hippocampal and cortical neurons under obesity conditions [
46]. GLP-1 improves neural differentiation through PI3K-AKT signaling in neuroblastoma cells [
47]. GLP-1 contributes to increased synaptic formation and memory consolidation in both diabetes and AD [
48]. GLP-1 and GLP-1 analogs prevent synaptic failure in AD [
49]. GLP-1 agonist injection enhances spatial, learning, and fear-related memory [
50]. According to our data and previous results, exendin-4, a GLP-1 agonist, prevents synaptic impairment and neural complexity in the brains of obese animal models with high levels of PA.
In conclusion, we corroborated the beneficial effects of adiponectin, BDNF, and GLP-1 on cortical neurons under PA toxicity. Adiponectin, BDNF, and GLP-1 prevent cortical neuronal death and maintain neural complexity and neuronal length in conditions of PA toxicity. We hypothesize that adiponectin, BDNF, and GLP-1 could be effective and safe options for treating metabolic imbalance–associated cognitive decline accompanied by synaptic failure.
Exercise could be an easy and effective approach to stimulate the secretion of adiponectin, BDNF, and GLP-1 in both nonobese and obese individuals. Several studies have shown that consistent exercise can increase the levels of adiponectin [
51], BDNF [
52], and GLP-1 [
53] in the blood of both nonobese and obese people.
NRF-2022R1A2C1006125
Chonnam National University Hwasun Hospitalhttps://doi.org/10.13039/501100019780
HCRI24023
NOTES
-
Funding: This study was supported by grants NRF-2022R1A2C1006125 (Juhyun Song) from the National Research Foundation of Korea (NRF) and HCRI24023 from the Chonnam National University Hwasun Hospital Institute for Biomedical Science, Korea (Juhyun Song).
-
Data Availability Statement: The data presented in this study are available within the article. The data supporting the findings of this study are available from the corresponding author upon reasonable request.
-
Conflict of Interest: The authors declare that they have no competing interests.
-
Author Contributions:
Conceptualization: Lee DH, Song J.
Formal analysis: Jo D, Ahn SY, Lee DH, Song J.
Funding acquisition: Lee DH, Song J.
Methodology: Jo D, Ahn SY, Choi SY, Choi Y, Lee DH, Song J.
Writing - review & editing: Lee DH, Song J.
ACKNOWLEDGEMENTS
REFERENCES
- 1. Engin A. The definition and prevalence of obesity and metabolic syndrome. Adv Exp Med Biol 2017;960:1-17.
- 2. Pan Y, Hui X, Hoo RL, Ye D, Chan CY, Feng T, Wang Y, Lam KS, Xu A. Adipocyte-secreted exosomal microRNA-34a inhibits M2 macrophage polarization to promote obesity-induced adipose inflammation. J Clin Invest 2019;129:834-849.
- 3. Benoit SC, Kemp CJ, Elias CF, Abplanalp W, Herman JP, Migrenne S, Lefevre AL, Cruciani-Guglielmacci C, Magnan C, Yu F, Niswender K, Irani BG, Holland WL, Clegg DJ. Palmitic acid mediates hypothalamic insulin resistance by altering PKC-theta subcellular localization in rodents. J Clin Invest 2009;119:2577-2589.
- 4. Amine H, Benomar Y, Taouis M. Palmitic acid promotes resistin-induced insulin resistance and inflammation in SH-SY5Y human neuroblastoma. Sci Rep 2021;11:5427.
- 5. Hernández-Cáceres MP, Toledo-Valenzuela L, Díaz-Castro F, Ávalos Y, Burgos P, Narro C, Peña-Oyarzun D, Espinoza-Caicedo J, Cifuentes-Araneda F, Navarro-Aguad F, Riquelme C, Troncoso R, Criollo A, Morselli E. Palmitic acid reduces the autophagic flux and insulin sensitivity through the activation of the free fatty acid receptor 1 (FFAR1) in the hypothalamic neuronal cell line N43/5. Front Endocrinol (Lausanne) 2019;10:176.
- 6. Melo HM, Seixas da Silva GD, Sant’Ana MR, Teixeira CV, Clarke JR, Miya Coreixas VS, de Melo BC, Fortuna JT, Forny-Germano L, Ledo JH, Oliveira MS, Figueiredo CP, Pardossi-Piquard R, Checler F, Delgado-García JM, Gruart A, Velloso LA, Balthazar ML, Cintra DE, Ferreira ST, De Felice FG. Palmitate is increased in the cerebrospinal fluid of humans with obesity and induces memory impairment in mice via pro-inflammatory TNF-α. Cell Reports 2020;30:2180-2194.e8.
- 7. Qiu T, Yang X, Wang J, Pan C, Chu X, Xiong J, Xie J, Chang Y, Wang C, Zhang J. Obesity-induced elevated palmitic acid promotes inflammation and glucose metabolism disorders through GPRs/NF-κB/KLF7 pathway. Nutr Diabetes 2022;12:23.
- 8. Jo D, Yoon G, Song J. Role of exendin-4 in brain insulin resistance, mitochondrial function, and neurite outgrowth in neurons under palmitic acid-induced oxidative stress. Antioxidants 2021;10:78.
- 9. Gariballa S, Alkaabi J, Yasin J, Al Essa A. Total adiponectin in overweight and obese subjects and its response to visceral fat loss. BMC Endocr Disord 2019;19:55.
- 10. Woodward L, Akoumianakis I, Antoniades C. Unravelling the adiponectin paradox: novel roles of adiponectin in the regulation of cardiovascular disease. Br J Pharmacol 2017;174:4007-4020.
- 11. Cisternas P, Martinez M, Ahima RS, William Wong G, Inestrosa NC. Modulation of glucose metabolism in hippocampal neurons by adiponectin and resistin. Mol Neurobiol 2019;56:3024-3037.
- 12. Waragai M, Adame A, Trinh I, Sekiyama K, Takamatsu Y, Une K, Masliah E, Hashimoto M. Possible involvement of adiponectin, the anti-diabetes molecule, in the pathogenesis of Alzheimer’s disease. J Alzheimers Dis 2016;52:1453-1459.
- 13. Dieni S, Matsumoto T, Dekkers M, Rauskolb S, Ionescu MS, Deogracias R, Gundelfinger ED, Kojima M, Nestel S, Frotscher M, Barde YA. BDNF and its pro-peptide are stored in presynaptic dense core vesicles in brain neurons. J Cell Biol 2012;196:775-788.
- 14. Sandrini L, Di Minno A, Amadio P, Ieraci A, Tremoli E, Barbieri SS. Association between obesity and circulating brain-derived neurotrophic factor (BDNF) levels: systematic review of literature and meta-analysis. Int J Mol Sci 2018;19:2281.
- 15. Park H, Poo MM. Neurotrophin regulation of neural circuit development and function. Nat Rev Neurosci 2013;14:7-23.
- 16. Antonsen KK, Klausen MK, Brunchmann AS, le Dous N, Jensen ME, Miskowiak KW, Fisher PM, Thomsen GK, Rindom H, Fahmy TP, Vollstaedt-Klein S, Benveniste H, Volkow ND, Becker U, Ekstrøm C, Knudsen GM, Vilsbøll T, Fink-Jensen A. Does glucagon-like peptide-1 (GLP-1) receptor agonist stimulation reduce alcohol intake in patients with alcohol dependence: study protocol of a randomised, double-blinded, placebo-controlled clinical trial. BMJ Open 2018;8:e019562.
- 17. Popoviciu MS, Păduraru L, Yahya G, Metwally K, Cavalu S. Emerging role of GLP-1 agonists in obesity: a comprehensive review of randomised controlled trials. Int J Mol Sci 2023;24:10449.
- 18. Nauck MA, Müller TD. Incretin hormones and type 2 diabetes. Diabetologia 2023;66:1780-1795.
- 19. Perry T, Lahiri DK, Sambamurti K, Chen D, Mattson MP, Egan JM, Greig NH. Glucagon-like peptide-1 decreases endogenous amyloid-beta peptide (Abeta) levels and protects hippocampal neurons from death induced by Abeta and iron. J Neurosci Res 2003;72:603-612.
- 20. Perry T, Lahiri DK, Chen D, Zhou J, Shaw KT, Egan JM, Greig NH. A novel neurotrophic property of glucagon-like peptide 1: a promoter of nerve growth factor-mediated differentiation in PC12 cells. J Pharmacol Exp Ther 2002;300:958-966.
- 21. Vrang N, Hansen M, Larsen PJ, Tang-Christensen M. Characterization of brainstem preproglucagon projections to the paraventricular and dorsomedial hypothalamic nuclei. Brain Res 2007;1149:118-126.
- 22. Chen XY, Chen L, Yang W, Xie AM. GLP-1 suppresses feeding behaviors and modulates neuronal electrophysiological properties in multiple brain regions. Front Mol Neurosci 2021;14:793004.
- 23. Williams DL, Lilly NA, Edwards IJ, Yao P, Richards JE, Trapp S. GLP-1 action in the mouse bed nucleus of the stria terminalis. Neuropharmacology 2018;131:83-95.
- 24. Yoon G, Kim YK, Song J. Glucagon-like peptide-1 suppresses neuroinflammation and improves neural structure. Pharmacol Res 2020;152:104615.
- 25. Hölscher C. Novel dual GLP-1/GIP receptor agonists show neuroprotective effects in Alzheimer’s and Parkinson’s disease models. Neuropharmacology 2018;136:251-259.
- 26. Zhang Y, Xue R, Zhang Z, Yang X, Shi H. Palmitic and linoleic acids induce ER stress and apoptosis in hepatoma cells. Lipids Health Dis 2012;11:1.
- 27. Bohlouli S, Khazaei M, Teshfam M, Hassanpour H. Adiponectin effect on the viability of human endometrial stromal cells and mRNA expression of adiponectin receptors. Int J Fertil Steril 2013;7:43-48.
- 28. Jourdi H, Kabbaj M. Acute BDNF treatment upregulates GluR1-SAP97 and GluR2-GRIP1 interactions: implications for sustained AMPA receptor expression. PLoS One 2013;8:e57124.
- 29. Hao S, Dey A, Yu X, Stranahan AM. Dietary obesity reversibly induces synaptic stripping by microglia and impairs hippocampal plasticity. Brain Behav Immun 2016;51:230-239.
- 30. Tsao TS, Lodish HF, Fruebis J. ACRP30, a new hormone controlling fat and glucose metabolism. Eur J Pharmacol 2002;440:213-221.
- 31. Kishore U, Reid KB. C1q: structure, function, and receptors. Immunopharmacology 2000;49:159-170.
- 32. Berg AH, Combs TP, Scherer PE. ACRP30/adiponectin: an adipokine regulating glucose and lipid metabolism. Trends Endocrinol Metab 2002;13:84-89.
- 33. Fry M, Smith PM, Hoyda TD, Duncan M, Ahima RS, Sharkey KA, Ferguson AV. Area postrema neurons are modulated by the adipocyte hormone adiponectin. J Neurosci 2006;26:9695-9702.
- 34. Ali T, Rehman SU, Khan A, Badshah H, Abid NB, Kim MW, Jo MH, Chung SS, Lee HG, Rutten BP, Kim MO. Adiponectin-mimetic novel nonapeptide rescues aberrant neuronal metabolic-associated memory deficits in Alzheimer’s disease. Mol Neurodegener 2021;16:23.
- 35. Pousti F, Ahmadi R, Mirahmadi F, Hosseinmardi N, Rohampour K. Adiponectin modulates synaptic plasticity in hippocampal dentate gyrus. Neurosci Lett 2018;662:227-232.
- 36. Shah SA, Yoon GH, Chung SS, Abid MN, Kim TH, Lee HY, Kim MO. Novel osmotin inhibits SREBP2 via the AdipoR1/AMPK/SIRT1 pathway to improve Alzheimer’s disease neuropathological deficits. Mol Psychiatry 2017;22:407-416.
- 37. Zhang D, Wang X, Wang B, Garza JC, Fang X, Wang J, Scherer PE, Brenner R, Zhang W, Lu XY. Adiponectin regulates contextual fear extinction and intrinsic excitability of dentate gyrus granule neurons through AdipoR2 receptors. Mol Psychiatry 2017;22:1044-1055.
- 38. Zhang D, Wang X, Lu XY. Adiponectin exerts neurotrophic effects on dendritic arborization, spinogenesis, and neurogenesis of the dentate gyrus of male mice. Endocrinology 2016;157:2853-2869.
- 39. Rauti R, Cellot G, D’Andrea P, Colliva A, Scaini D, Tongiorgi E, Ballerini L. BDNF impact on synaptic dynamics: extra or intracellular long-term release differently regulates cultured hippocampal synapses. Mol Brain 2020;13:43.
- 40. Sun Y, Lim Y, Li F, Liu S, Lu JJ, Haberberger R, Zhong JH, Zhou XF. ProBDNF collapses neurite outgrowth of primary neurons by activating RhoA. PLoS One 2012;7:e35883.
- 41. Christel CM, Denardo DF. Absence of exendin-4 effects on postprandial glucose and lipids in the Gila monster, Heloderma suspectum
. J Comp Physiol B 2007;177:129-134.
- 42. De León DD, Crutchlow MF, Ham JY, Stoffers DA. Role of glucagon-like peptide-1 in the pathogenesis and treatment of diabetes mellitus. Int J Biochem Cell Biol 2006;38:845-859.
- 43. Zago AM, Carvalho FB, Rahmeier FL, Santin M, Guimaraes GR, Gutierres JM, da C Fernandes M. Exendin-4 prevents memory loss and neuronal death in rats with sporadic Alzheimer-like disease. Mol Neurobiol 2024;61:2631-2652.
- 44. Gumuslu E, Mutlu O, Celikyurt IK, Ulak G, Akar F, Erden F, Ertan M. Exenatide enhances cognitive performance and upregulates neurotrophic factor gene expression levels in diabetic mice. Fundam Clin Pharmacol 2016;30:376-384.
- 45. Zhao F, Li J, Wang R, Xu H, Ma K, Kong X, Sun Z, Niu X, Jiang J, Liu B, Li B, Duan F, Chen X. Exendin-4 promotes actin cytoskeleton rearrangement and protects cells from Nogo-A-Δ20 mediated spreading inhibition and growth cone collapse by down-regulating RhoA expression and activation via the PI3K pathway. Biomed Pharmacother 2019;109:135-143.
- 46. Wang M, Yoon G, Song J, Jo J. Exendin-4 improves long-term potentiation and neuronal dendritic growth in vivo and in vitro obesity condition. Sci Rep 2021;11:8326.
- 47. Yang JL, Lin YT, Chen WY, Yang YR, Sun SF, Chen SD. The neurotrophic function of glucagon-like peptide-1 promotes human neuroblastoma differentiation via the PI3K-AKT axis. Biology (Basel) 2020;9:348.
- 48. Zheng J, Xie Y, Ren L, Qi L, Wu L, Pan X, Zhou J, Chen Z, Liu L. GLP-1 improves the supportive ability of astrocytes to neurons by promoting aerobic glycolysis in Alzheimer’s disease. Mol Metab 2021;47:101180.
- 49. Gault VA, Hölscher C. GLP-1 agonists facilitate hippocampal LTP and reverse the impairment of LTP induced by beta-amyloid. Eur J Pharmacol 2008;587:112-117.
- 50. During MJ, Cao L, Zuzga DS, Francis JS, Fitzsimons HL, Jiao X, Bland RJ, Klugmann M, Banks WA, Drucker DJ, Haile CN. Glucagon-like peptide-1 receptor is involved in learning and neuroprotection. Nat Med 2003;9:1173-1179.
- 51. Lin D, Sturgeon KM, Gordon BR, Brown JC, Sears DD, Sarwer DB, Schmitz KH. WISER survivor trial: combined effect of exercise and weight loss interventions on adiponectin and leptin levels in breast cancer survivors with overweight or obesity. Nutrients 2023;15:3453.
- 52. Ribeiro D, Petrigna L, Pereira FC, Muscella A, Bianco A, Tavares P. The impact of physical exercise on the circulating levels of BDNF and NT 4/5: a review. Int J Mol Sci 2021;22:22.
- 53. Sandsdal RM, Juhl CR, Jensen SB, Lundgren JR, Janus C, Blond MB, Rosenkilde M, Bogh AF, Gliemann L, Jensen JB, Antoniades C, Stallknecht BM, Holst JJ, Madsbad S, Torekov SS. Combination of exercise and GLP-1 receptor agonist treatment reduces severity of metabolic syndrome, abdominal obesity, and inflammation: a randomized controlled trial. Cardiovasc Diabetol 2023;22:41.